Dobzhansky, T., (1951). Genetics and the Origin of Species, 3.ed, New York: Columbia University Press |
Silverman M. og Simon M., (1983) “Phase variation and related systems” in Shaphiro J.A. (red), (1983). Mobile genetic elements, new York: Ac. Press, ss.537-557 |
Darnell et al (1986). Molecular Cell Biology. New York: Scientific American Books. |
Griffits et al (1993). An Introduction to Genetic Analysis, New York: Freeman |
McClintock (1978) “Mechanisms that rapidly reorganise the genome” Reprinted in Frederik and Botstein (1992) |
Cairns et al (1988). “The Origin of mutants” Nature vol. 335, s.142-145
|
Stahl F.W. (1988) “A unicorn in the garden,” Nature vol. 335, ss.112-113
|
Benson S.A. (1988). “Is bacterial evolution random or selective?” Nature vol. 336, ss21-22 |
Sniegowski P.D. & Lenski R.E.(1995). “Mutation and adaptation: The directed mutation controversy in evolutionary perspective.” Annual Review of Ecology and Systematics vol. 26. ss.553-578 |
Foster P.L. og Trimarchi J.M. (1994). “Adaptive reversion of a frameshift mutation in Escherichia coli in simple base deletions in homopolymeric runs,” Science, vol.265, ss.407-409 |
Rosenberg S.M. et al. (1994). “Adaptive mutations by deletions in small mononucleotide Repeats, Science vol. 265, ss.405-407 |
Radicella et al. (1995). “Adaptive mutation in Escherichia coli: A Role for Conjugation,” Science vol. 268, ss.418-420 |
Shaphiro J.A. (1995) “Adaptive mutation: Who’s really in the garden?” Science vol. 268, ss.373-374 |
Holiday R. (1989). “A different kind of inheritance,” Scientific Am. Vol.260 June |
Herskowitz og Hagen (1980). “The lysis-lysogeny decision of phage lambda: explicit programming on responsiveness.” Annual Review of Genetics, vo14, ss.399-446 |
Johnston T.D. og Gottlieb (1990), “Neophenogenesis: A developmental theory of phenotype evolution,” Journal of Theoretical Biology, vol.147, ss.471-495 |
Kalat J.W. (1985) In Johnston and Pietrewicz (1985), ss. 119-141 |
Galef B.G. (1985) ) In Johnston and Pietrewicz (1985), ss.143-146 |
Fryer, Greenwood og Peak (1983, 1985) “Punctuated equilibria, morphological stasis and the paleontological documentation of speciation: a biological appraisal of a case history in an African lake,” Biological Journal of the Linnean Society, vol. 20, ss.195-205 |
McDonald J.F. (1990), "Macroevolution and retroviral elements," BioScience, vol.40, ss183-191 |
Wilson et al. (1974). "Two types of molecular evolution. Evidences from studies of interspecific hybridization," Proceedings National Academy of Science USA, Vol.71, pp.2843-2847 |
West-Eberhard (1989). "Phenotype plasticity and the origins of diversity," Annual Review of Ecological Systems, vo.20, ss.249-278 |
Schlichting C.D. (1986). “The evolution of phenotype placticity in plants,” Ann. Review of Ecological Systems, vol. 17. ss.667-693 |
Stearns S.C. (1989). “The evolutionary significanse of phenotype plasticity,” BioScience, vol.39, ss.436-445 |
Bradshaw A.D. (1965). “Evolutionary significanse of phenotype plasticity in plants,” Advances in Genetics, vol.13, ss.115-155 |
Stearns SS(1989), The evolutionary significanse of phenotype plasticity,” Bioscience vol. 39, ss.436-445 |
Ho og Saunders, (1979), "Beyond Neo-Darwinism, An epigenetic approach to evolution," Journal of Theoretical Biology, vo.78. s.573-591 |
Conant, S. (1988). “Saving endangered species by translocation.” BioScience, vol.38, ss.254-257 |
Pimm S.L. (1988). “Rapid morhological change in an introduced bird,” Trends in Evolution and Ecology, vol.3. ss.290-291 |
Reznick D. et al. (1997). “Evalutation of the rate of evolution in natural populations of guppies (Poecilia reticulate). Science, vol. 275, ss.1934-1937 |
Packard A. (1972). “Cecalophods and fish: The limits of convergence,” Biological Review, vol.47, ss.241-307 |
Collier B.D. et al (1973) Dynamic Ecology, Englewood Cliffs: Prentice Hall |
Sumner F.B. (1909). “Some effects of external conditions on the white mouse,” Journal of Experimental Zoology, vol.7, ss.97-155 |
Schreider E.(1964). “Ecological rules, body-heat regulation, and human evolution,” Ecology, vol.18, ss.1-9 |
Brooks J.L. (1957) In Mayr (1957), sss.81-123 |
Murray C.B. og Beacham T.D. (1989). “Responses in meristic characters in chum salmon (Onchorhynchus keta) to temperature changes during development,” Canadian Journal of Zoology, vol.67, ss.596-600 |
Stoffutvalg og bilder ved Asbjørn E. Lund {evt. kommentar i klammeparentes}
.
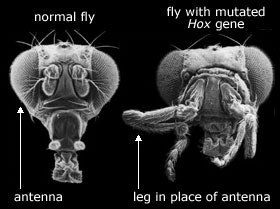 Bill McClintock, som mottok Nobelprisen i 1983 for arbeid med genetiske rearrangement, bemerket at det er indikasjoner på at disse genetiske modifikasjonene inntreffer som respons på stress (McClintock 1978). Vi vet ikke ennå hva normalfunksjonen er, bortsett fra å spre bakteriell resistens. B. Wanner ved Emory universitet har foreslått av genetisk rearrangement kan være del av ett kontrollsystem i bakterier som produserer arvelige endringer som motsvar på signaler fra omgivelsene (Wanner 1985). V. Walbot ved Stanford universitet og Chr. Cullis ved Case Western Reserve universitet, har bemerket at hvert genetisk rearrangement kan være ansvarlig for raske genetiske endringer, særlig i planter (Walbot og Cullis 1985). Ecoli kan vanligvis ernære seg på melkesukker (laktose) fordi det har en streng av enzymer som benytte det i stoffskiftet. For å teste ut preparerte B. Hall en genstreng som manglet det som kodet for det 1. enzymet i strengen. Bakteriestrengen kunne dermed ikke leve på melkesukker. Men da disse bakteriene ernærte seg på en annen næringskilde, ble to mutasjoner oppdaget. Den ene var i et hittil ukjent strukturelt gen, og den andre i kontrollgenet. Fordi det muterte strukturelle genet kodet for et enzym som kan utføre savnede 1.steg i laktose-stoffskiftet, var det i stand til å ernære seg på melkesukker. Det muterte genet hadde vært til stede hele tiden, men var inaktivt. Den normale funksjonen ved det er ukjent. Hall kalte det et 'kryptisk gen'.
Bill McClintock, som mottok Nobelprisen i 1983 for arbeid med genetiske rearrangement, bemerket at det er indikasjoner på at disse genetiske modifikasjonene inntreffer som respons på stress (McClintock 1978). Vi vet ikke ennå hva normalfunksjonen er, bortsett fra å spre bakteriell resistens. B. Wanner ved Emory universitet har foreslått av genetisk rearrangement kan være del av ett kontrollsystem i bakterier som produserer arvelige endringer som motsvar på signaler fra omgivelsene (Wanner 1985). V. Walbot ved Stanford universitet og Chr. Cullis ved Case Western Reserve universitet, har bemerket at hvert genetisk rearrangement kan være ansvarlig for raske genetiske endringer, særlig i planter (Walbot og Cullis 1985). Ecoli kan vanligvis ernære seg på melkesukker (laktose) fordi det har en streng av enzymer som benytte det i stoffskiftet. For å teste ut preparerte B. Hall en genstreng som manglet det som kodet for det 1. enzymet i strengen. Bakteriestrengen kunne dermed ikke leve på melkesukker. Men da disse bakteriene ernærte seg på en annen næringskilde, ble to mutasjoner oppdaget. Den ene var i et hittil ukjent strukturelt gen, og den andre i kontrollgenet. Fordi det muterte strukturelle genet kodet for et enzym som kan utføre savnede 1.steg i laktose-stoffskiftet, var det i stand til å ernære seg på melkesukker. Det muterte genet hadde vært til stede hele tiden, men var inaktivt. Den normale funksjonen ved det er ukjent. Hall kalte det et 'kryptisk gen'. mutasjonene, og kunne ernære seg på salicin. I fravær av salicin var slettingshastigheten av IS103 for lav til å måles, mens i nærvær av det hadde 89% av cellene i en koloni gjennomgått slettingen av IS103.
mutasjonene, og kunne ernære seg på salicin. I fravær av salicin var slettingshastigheten av IS103 for lav til å måles, mens i nærvær av det hadde 89% av cellene i en koloni gjennomgått slettingen av IS103. Genetikere har forsøkt å teste eksperimentet nevnt ovenfor. Testingen har ikke løst striden, men heller utvidet den. Noen er mot å trekke vidtgående konklusjoner av forsøket (Sniegowski & Lenski 1995) og mener resultatet ikke er klarlagt ennå, mens andre støtter at effektene i forsøket er reelle (Foster 1992). Flere nylige forsøk har vist en annen slag mutasjoner som synes å være tilpassede og frembragt av omgivelsene. Disse viser at i nærvær av laktose vil tilpassede mutasjoner aktivere et hvilende gen, som koder for et enzym som vil spalte laktose i E.Coli, være av et annet slag enn de som inntreffer i fravær av laktose (Foster og Trimarchi 1994; Rosenberg et al. 1994, Gahtski og Roth 1995, Radicelle et al. 1995), samt Culotta (1994) og Shaphiro (1995). Darwinistiske evolusjonister ser den ikke-tilfeldige tolkningen av disse eksperimentelle resultatene som åpenbart feilaktige, fordi de motsier det NDT-dogme. Forfatteren (Dr Lee Spetner) ser denne tolkningen som en bekreftelse på bakterielt nivå av ikke-tilfeldig variasjon, som er indikert av mange eks. i planter og dyr. Dette er eks. som darwinistiske evolusjonister for det meste har ignorert, fordi den ikke 'passer inn'. Motstand mot fortolkning om ikke-tilfeldig variasjon, skyldes at en nekter å forlate darwinistisk agenda om at livet oppsto og utviklet seg spontant. Med en slik agenda, så er ikke-tilfeldig tilpasset variasjon, som oppstår ved at signaler fra omgivelsene slår PÅ ett allerede eksisterende sett av gener, vanskelig å gjøre rede for. Tendensen en har til å akseptere ikke-tilfeldig fortolkning av eksperimentene på, avhenger av hvor sterkt en insisterer på den darwinistiske agendaen. Hvilken fortolkning som er korrekt, vil til slutt bli avgjort i laboratoriene.
Genetikere har forsøkt å teste eksperimentet nevnt ovenfor. Testingen har ikke løst striden, men heller utvidet den. Noen er mot å trekke vidtgående konklusjoner av forsøket (Sniegowski & Lenski 1995) og mener resultatet ikke er klarlagt ennå, mens andre støtter at effektene i forsøket er reelle (Foster 1992). Flere nylige forsøk har vist en annen slag mutasjoner som synes å være tilpassede og frembragt av omgivelsene. Disse viser at i nærvær av laktose vil tilpassede mutasjoner aktivere et hvilende gen, som koder for et enzym som vil spalte laktose i E.Coli, være av et annet slag enn de som inntreffer i fravær av laktose (Foster og Trimarchi 1994; Rosenberg et al. 1994, Gahtski og Roth 1995, Radicelle et al. 1995), samt Culotta (1994) og Shaphiro (1995). Darwinistiske evolusjonister ser den ikke-tilfeldige tolkningen av disse eksperimentelle resultatene som åpenbart feilaktige, fordi de motsier det NDT-dogme. Forfatteren (Dr Lee Spetner) ser denne tolkningen som en bekreftelse på bakterielt nivå av ikke-tilfeldig variasjon, som er indikert av mange eks. i planter og dyr. Dette er eks. som darwinistiske evolusjonister for det meste har ignorert, fordi den ikke 'passer inn'. Motstand mot fortolkning om ikke-tilfeldig variasjon, skyldes at en nekter å forlate darwinistisk agenda om at livet oppsto og utviklet seg spontant. Med en slik agenda, så er ikke-tilfeldig tilpasset variasjon, som oppstår ved at signaler fra omgivelsene slår PÅ ett allerede eksisterende sett av gener, vanskelig å gjøre rede for. Tendensen en har til å akseptere ikke-tilfeldig fortolkning av eksperimentene på, avhenger av hvor sterkt en insisterer på den darwinistiske agendaen. Hvilken fortolkning som er korrekt, vil til slutt bli avgjort i laboratoriene. Bilde 6
Bilde 6
 I følge det som er beskrevet her, kunne en vente at selv større evolusjonære endringer kan bære med seg kun mindre genetiske endringer. Om de skyldes den andre type av genetisk variasjon, nevnt ovenfor, så ville det ikke være noen endring i det hele i DNA-sekvensen. Om makroevolusjon skjedde i følge NDT, så ville store evolusjonære endringer følges av store DNA-endringer. John McDonald har pekt på mangel av korrelasjon mellom størrelser av fenotype-endringer og DNA-endringer. Forskjeller i DNA mellom arter synes ikke å være relatert til deres antatte evolusjonære divergens (McDonald 1990). I det han siterer arbeidet til Alan Wilson og medarbeidere (Wilson et al 1974), bemerket McDonald forskjellene mellom frosk og pattedyr i så måte. Det finnes to froskeslag som er meget like, men skiller seg mye i genomet. Pattedyrene har til tross for store ulikheter i fenotypen, lite forskjeller i genotypen. Disse dataene indikerer at størrelsen på genetiske endringer ikke trenger være relatert til graden av fenotype endringer. Faktisk kan mye genetisk endring være irrelevant for evolusjonær endring.
I følge det som er beskrevet her, kunne en vente at selv større evolusjonære endringer kan bære med seg kun mindre genetiske endringer. Om de skyldes den andre type av genetisk variasjon, nevnt ovenfor, så ville det ikke være noen endring i det hele i DNA-sekvensen. Om makroevolusjon skjedde i følge NDT, så ville store evolusjonære endringer følges av store DNA-endringer. John McDonald har pekt på mangel av korrelasjon mellom størrelser av fenotype-endringer og DNA-endringer. Forskjeller i DNA mellom arter synes ikke å være relatert til deres antatte evolusjonære divergens (McDonald 1990). I det han siterer arbeidet til Alan Wilson og medarbeidere (Wilson et al 1974), bemerket McDonald forskjellene mellom frosk og pattedyr i så måte. Det finnes to froskeslag som er meget like, men skiller seg mye i genomet. Pattedyrene har til tross for store ulikheter i fenotypen, lite forskjeller i genotypen. Disse dataene indikerer at størrelsen på genetiske endringer ikke trenger være relatert til graden av fenotype endringer. Faktisk kan mye genetisk endring være irrelevant for evolusjonær endring.
 Krabber lever av snegler, men kan ikke spise snegler med tykt skall. Snegler kan på ett eller annet vis vite om krabber er i området. I nærvær av krabber får de tykkere skall (Stearns 1989). Denne tilpasningen hjelper klart å beskytte snegler fra krabber. Snegler er selv rovdyr som lever av et dyr i småkrepsgruppen (rankefoting-barnacle). Når disse senser snegler beskytter de seg ved å vokse inn i en krumbøyd stilling som hindrer sneglene å spise dem. Når det ikke finnes snegler omkring, går rankefotingen tilbake til sin opprinnelige form (Stearns 1986). Det forfatteren foreslår her, er at organismer har en innebygd evne til å tilpasse seg omgivelsene. Han foreslår at i den grad evolusjon foregår, foregår det på individnivå hos organismen. Dette forslaget skiller seg skarpt fra tesen til NDT, som holder på at evolusjon inntreffer bare på populasjonsnivå. Organismer inneholder informasjonen som setter dem i stand til å utvikle en fenotype tilpasset en variasjon av omgivelser. Tilpasningen kan inntreffe ved en endring i genomet gjennom en genetisk endring trigget av omgivelsene, eller det kan inntreffe uten noen genetisk endring.
Krabber lever av snegler, men kan ikke spise snegler med tykt skall. Snegler kan på ett eller annet vis vite om krabber er i området. I nærvær av krabber får de tykkere skall (Stearns 1989). Denne tilpasningen hjelper klart å beskytte snegler fra krabber. Snegler er selv rovdyr som lever av et dyr i småkrepsgruppen (rankefoting-barnacle). Når disse senser snegler beskytter de seg ved å vokse inn i en krumbøyd stilling som hindrer sneglene å spise dem. Når det ikke finnes snegler omkring, går rankefotingen tilbake til sin opprinnelige form (Stearns 1986). Det forfatteren foreslår her, er at organismer har en innebygd evne til å tilpasse seg omgivelsene. Han foreslår at i den grad evolusjon foregår, foregår det på individnivå hos organismen. Dette forslaget skiller seg skarpt fra tesen til NDT, som holder på at evolusjon inntreffer bare på populasjonsnivå. Organismer inneholder informasjonen som setter dem i stand til å utvikle en fenotype tilpasset en variasjon av omgivelser. Tilpasningen kan inntreffe ved en endring i genomet gjennom en genetisk endring trigget av omgivelsene, eller det kan inntreffe uten noen genetisk endring. Finker, f.eks. i form av spurver og sangere, er den største, mest sette og diversifiserte fuglefamilie. De utgjøre den største familien (Fringillidae) av den største orden (Paseriformes) i klassen fugler. Deres evolusjon er blitt mer studert enn for noen annen fuglefamilie. Darwins undersøkelse av finker på Galapagos er velkjent, han fant 13 ulike finkearter hver spesialisert i hver sin nisje. Formen og styrken i nebbene -hvorvidt de er spisse og lange, eller brede og kraftige, så vel som musklene er tilpasset typen mat de spiser. Spørsmålet blir hvordan de ulike finkene oppsto og hvorfor de skiller seg fra andre finker? De fleste eksperter holder med Darwins hypotese, modifisert gjennom NDT. Hypotesen er basert på formodningen at små tilfeldige mutasjoner kan ha ført til tilpassede endringer. Noen vitenskapsfolk mener ikke det: Ho og Saunders har uttalt: "Det strekker troverdigheten å forestille seg, f.eks., at hakkespetten først fikk en langt nebb av noen tilfeldige mutasjoner, fulgt av andre mutasjoner som fikk den til å lete etter larver i barken på trærne." Videre er det ikke beviser for det, og det strider mot
Finker, f.eks. i form av spurver og sangere, er den største, mest sette og diversifiserte fuglefamilie. De utgjøre den største familien (Fringillidae) av den største orden (Paseriformes) i klassen fugler. Deres evolusjon er blitt mer studert enn for noen annen fuglefamilie. Darwins undersøkelse av finker på Galapagos er velkjent, han fant 13 ulike finkearter hver spesialisert i hver sin nisje. Formen og styrken i nebbene -hvorvidt de er spisse og lange, eller brede og kraftige, så vel som musklene er tilpasset typen mat de spiser. Spørsmålet blir hvordan de ulike finkene oppsto og hvorfor de skiller seg fra andre finker? De fleste eksperter holder med Darwins hypotese, modifisert gjennom NDT. Hypotesen er basert på formodningen at små tilfeldige mutasjoner kan ha ført til tilpassede endringer. Noen vitenskapsfolk mener ikke det: Ho og Saunders har uttalt: "Det strekker troverdigheten å forestille seg, f.eks., at hakkespetten først fikk en langt nebb av noen tilfeldige mutasjoner, fulgt av andre mutasjoner som fikk den til å lete etter larver i barken på trærne." Videre er det ikke beviser for det, og det strider mot  Thomas Smith ved Berkeley universitet i California, studerte afrikanske finker i Kamerun. Han fant at finkene kunne få avkom med to ulike nebbstørrelser. Han sa ikke hvorvidt miljøet spilte noen rolle i hvilken, men han rapporterte at nebbstørrelsene var tilpassede, hver i sin egen nisje. Fuglene med større nebb knekker lett harde frø, mens de med små nebb ernærer seg mer effektivt på myke frø, enn de med store nebb. Disse resultatene viser at nebbstørrelsen i finker kan endre seg fra en tilpasset type til en annen med diett. David Reznick og teamet hans ved California universitet har studert guppy-fiskearter i naturen. De endrer formeringstidspunkt og kvantum ut fra omgivelsene. Om det finnes rovfisk (killfish) som jakter på fiskeyngel, så har guppiene en tendens til å formere seg sent og få færre avkom. Det finnes en rovfisk av art alta, som jakter på større, voksne guppier. Normalt vil guppier som lever i lag med alta-rovfisken modnes tidligere og få flere og mindre avkom enn de som lever i lag med killfish. Dette er tilfelle i Aripo elva i Trinidad, der guppier lever sammen med alta-ciklider. Reznick og teamet hans tok 200 guppier fra Aripo-elven, og plasserte dem i ei sideelv der det finnes killfish, men ingen ciklider. Det dukket snart opp endringer i den ny-introduserte bestanden av guppier. Fiskepopulasjonen endret seg snart til det som er vanlig for guppier som lever i lag med killfish, og Reznick fant at endringen var arvelig. Endringen i guppy-bestanden ble observert så snart noen av de første fiskene ble fanget, som var etter bare 2 år. Ett trekk de studerte, modenhetsalder for fisken, nådde sitt termineringsverdi etter bare 4 år. Evolusjonsraten som kan kalkuleres ut fra observasjonen, er ca. 10 millioner ganger endringsraten ved observasjoner fra fossilarkivet (Reznick et al. 1997) Reznick fortolket resultatene som at naturlig seleksjon virket på variasjon allerede i populasjonen. En mer fornuftig forklaring er at nærvær av rovfisken induserte endringer i individuelle fiskene.
Thomas Smith ved Berkeley universitet i California, studerte afrikanske finker i Kamerun. Han fant at finkene kunne få avkom med to ulike nebbstørrelser. Han sa ikke hvorvidt miljøet spilte noen rolle i hvilken, men han rapporterte at nebbstørrelsene var tilpassede, hver i sin egen nisje. Fuglene med større nebb knekker lett harde frø, mens de med små nebb ernærer seg mer effektivt på myke frø, enn de med store nebb. Disse resultatene viser at nebbstørrelsen i finker kan endre seg fra en tilpasset type til en annen med diett. David Reznick og teamet hans ved California universitet har studert guppy-fiskearter i naturen. De endrer formeringstidspunkt og kvantum ut fra omgivelsene. Om det finnes rovfisk (killfish) som jakter på fiskeyngel, så har guppiene en tendens til å formere seg sent og få færre avkom. Det finnes en rovfisk av art alta, som jakter på større, voksne guppier. Normalt vil guppier som lever i lag med alta-rovfisken modnes tidligere og få flere og mindre avkom enn de som lever i lag med killfish. Dette er tilfelle i Aripo elva i Trinidad, der guppier lever sammen med alta-ciklider. Reznick og teamet hans tok 200 guppier fra Aripo-elven, og plasserte dem i ei sideelv der det finnes killfish, men ingen ciklider. Det dukket snart opp endringer i den ny-introduserte bestanden av guppier. Fiskepopulasjonen endret seg snart til det som er vanlig for guppier som lever i lag med killfish, og Reznick fant at endringen var arvelig. Endringen i guppy-bestanden ble observert så snart noen av de første fiskene ble fanget, som var etter bare 2 år. Ett trekk de studerte, modenhetsalder for fisken, nådde sitt termineringsverdi etter bare 4 år. Evolusjonsraten som kan kalkuleres ut fra observasjonen, er ca. 10 millioner ganger endringsraten ved observasjoner fra fossilarkivet (Reznick et al. 1997) Reznick fortolket resultatene som at naturlig seleksjon virket på variasjon allerede i populasjonen. En mer fornuftig forklaring er at nærvær av rovfisken induserte endringer i individuelle fiskene. La oss kalle hypotesen som foreslås i dette kapitlet for En Ikke-Tilfeldig Evolusjons Hypotese, for korthets skyld: EITEH. Vi har sett tidligere at NDT ikke kan gjøre rede for de mange såkalte konvergenser i levende organismer. Hva som kan ha virket som konvergens, kan imidlertid forståes ifølge EITEH å være resultat av at omgivelsene direkte influerer fenotypen evt. genotypen. Det som i følge NDT er konvergens, kan faktisk være ulike arters lignende innebygde tilsvar på samme signaler fra omgivelsene. I følge EITEH er det ingen overraskelse at en finner to grupper av organismer i atskilte klasser, som har lignende tilpassede trekk i lignende omgivelser. For drøyt 90 år siden skrev Göthe Turesson [1925]: " Det har lenge vært kjent at planter, som er fjernt fra hverandre i slektskap, men som bebor samme tilholdssted, også kan vise lignende eller samme trekk i vekst-former." Fisk og blekksprut er ulike dyrerekker, likevel når de okkuperer samme nisje i omgivelsene så kan de ha flere trekk felles. Det kan være fellestrekk i synssystem, bevegelsessystem, opplysningssystem. Fisk i ulike områder kan variere seg i mellom på disse egenskapene, slik blekkspruter også gjør. Men når de to rekkene deler samme nisje, så er det også funnet felles trekk (Packard 1972). Det er mange eks. på at artene deler felles plastiske trekk. Det som synes som konvergens, kan bare være plastisk respons på samme omgivelser.
La oss kalle hypotesen som foreslås i dette kapitlet for En Ikke-Tilfeldig Evolusjons Hypotese, for korthets skyld: EITEH. Vi har sett tidligere at NDT ikke kan gjøre rede for de mange såkalte konvergenser i levende organismer. Hva som kan ha virket som konvergens, kan imidlertid forståes ifølge EITEH å være resultat av at omgivelsene direkte influerer fenotypen evt. genotypen. Det som i følge NDT er konvergens, kan faktisk være ulike arters lignende innebygde tilsvar på samme signaler fra omgivelsene. I følge EITEH er det ingen overraskelse at en finner to grupper av organismer i atskilte klasser, som har lignende tilpassede trekk i lignende omgivelser. For drøyt 90 år siden skrev Göthe Turesson [1925]: " Det har lenge vært kjent at planter, som er fjernt fra hverandre i slektskap, men som bebor samme tilholdssted, også kan vise lignende eller samme trekk i vekst-former." Fisk og blekksprut er ulike dyrerekker, likevel når de okkuperer samme nisje i omgivelsene så kan de ha flere trekk felles. Det kan være fellestrekk i synssystem, bevegelsessystem, opplysningssystem. Fisk i ulike områder kan variere seg i mellom på disse egenskapene, slik blekkspruter også gjør. Men når de to rekkene deler samme nisje, så er det også funnet felles trekk (Packard 1972). Det er mange eks. på at artene deler felles plastiske trekk. Det som synes som konvergens, kan bare være plastisk respons på samme omgivelser.  Jordans regel: Mange fiskearter synes å ha større ryggvirvler når de lever i kaldt vann, enn de samme artene har i varmt vann (Screider 1964). Disse ulikhetene har vært vist å avhenge av temperaturen fiskene er vokst opp i. (Brooks 1957; Hubbs 1922, 1926; Murray og Beacham 1989; Johnston og Gottlieb 1990).
Jordans regel: Mange fiskearter synes å ha større ryggvirvler når de lever i kaldt vann, enn de samme artene har i varmt vann (Screider 1964). Disse ulikhetene har vært vist å avhenge av temperaturen fiskene er vokst opp i. (Brooks 1957; Hubbs 1922, 1926; Murray og Beacham 1989; Johnston og Gottlieb 1990).